Возбудимость сердечной мышцы неодинакова во всех участках сердца. Наиболее возбудимым синусоатриальным узел. Возбудимость пучка Гиса значительно меньше. Хотя во время сокращения мышца сердца возбудима. Но в этот период, который почти совпадает с систолой, самые сильные искусственные раздражения сердца не вызывают нового сокращения вследствие «конфликта двух сильных возбуждений, слишком близко поставленных одно к другому во времени в одном и том же субстрате» (А. А. Ухтомский). Это состояние полное потери возбудимости во время сокращения сердца обозначается как абсолютная рефрактерность. После этого во время расслабления мышцы сердца при раздражении сердца ударом индукционного электрического тока, вследствие изменения интервала времени между двумя возбуждениями и изменения функционального состояния сердца, может быть получено вне очереди, но более слабое сокращение.
Этот второй период неполной возбудимости во время расслабления сердца обозначается как относительная рефрактерность. Непосредственно после периода относительной рефрактерности наблюдается кратковременное повышение возбудимости - экзальтационная фаза. Продолжительность абсолютной и относительной рефрактерности зависит от длительности- сердечного цикла. Период абсолютной рефрактерности синусоатриального узла у человека доходит до 0,3 с., предсердий — от 0,06 до 0,12 с., а желудочков - от 0,3 до 0,4 с.
Благодаря продолжительной рефрактерности сердце отвечает на продолжительное раздражение ритмическими сокращениями и в обычных условиях на может прийти в состояние тетануса.
Если на желудочек сердца холоднокровного животного нанести раздражение до прихода очередного автоматического импульса, т. е. в периоде относительной рефрактерности, то возникает преждевременное сокращение сердца - экстрасистола, за которым следует компенсаторная пауза, по продолжительности превышающая обычную.
Экстрасистолы возникают при изменениях в проводящей системе или в мышце сердца. Влияние на изменение возбудимости обозначается как батмотропное.
Сокращение сердечной мышцы не усиливается с увеличением раздражения. Если непосредственно наносить раздражение на сердечную мышцу, наращивая каждый раз величину раздражения, то обнаруживается следующий факт. Вначале при слабых раздражениях мышца па них не реагирует сокращением, затем при некотором повышении величины раздражения она сокращается. Это сокращение максимальное. Дальнейшее увеличение силы раздражения уже не увеличивает сокращения сердечной мышцы (Г. Боудич, 1871).
Однако это только частный случай, а не правило, так как высота сокращения мышцы сердца («все») изменяется и зависит, от ее возбудимости и лабильности, т. е. от ее функционального состояния. «Ничего» также не существует, так как при подпороговых раздражениях возникает возбуждение, которое суммируется при определенной частоте раздражений.
Величина наибольшего сокращения сердечной мышцы зависит от уровня обмена веществ в ней. Влияние на силу сердечных сокращений обозначается как инотропное.
В процессе филогенеза выработалась способность сердечной мышцы увеличивать силу своих сокращений в зависимости от увеличения количества притекающей к сердцу крови и повышения давления крови в артериальной системе.
Увеличение притока крови к сердцу и повышение кровяного давления в физиологических условиях вызываются мышечной работой и некоторыми эмоциями.
Как увеличивает сердце силу своих сокращений при повышенных нагрузках?
Сила сокращений сердца увеличивается благодаря увеличению начальной длины мышечных волокон (Старлинг, 1916).
Мышечные волокна имеют определенную длину при диастоле сердца во время покоя организма, перед началом сокращения сердца (начальная длина). При увеличении притока крови к сердцу и при затруднениях оттока, вызванных повышением кровяного давления, сердце в диастоле от переполнения полости кровью растягивается сильнее, следовательно, начальная длина мышечных волокон сердца увеличивается. Чем больше приток крови к сердцу или чем больше кровяное давление, затрудняющее отток крови, тем больше начальное растяжение мышечных волокон.
На изолированных мышцах установлено, что сокращении скелетной и сердечной мышц прямо пропорциональна начальниц длине мышечных волокон. Чем больше начальная длина волокон, тем сильнее сокращение. Поэтому при увеличении начальной длины волокон сердца оно сильнее сокращается во время систолы и благодаря этому увеличивается количество выбрасываемой .
Большое значение имеет кровоснабжение и питание сердечной мышцы. Чем лучше питание мышцы, тем меньше она предварительно растягивается.
В естественных условиях при отсутствии дополнительного растяжения сердца увеличение сокращений - результат усиления в сердечном мускуле под влиянием нервной системы (трофическое влияние).
Когда сердечная мышца утомляется, то в сердце падает и оно растягивается. Способность сердца производить прежнюю работу при утомлении зависит от степени растяжения его мышечных волокон.
Степень растяжения сердца определяется толщиной и состоянием сердечной мышцы. Максимально сердце может расшириться до перикарда, который, таким образом, обусловливает предельное расширение сердца.
К основным свойствам сердечной мышцы относятся: 1) автоматия, 2) возбудимость, 3) проводимость и 4) сократимость.
АВТОМАТИЯ
Способность к ритмическому сокращению без всяких видимых раздражений под влиянием импульсов, возникающих в самом органе, является характерной особенностью сердца. Это свойство называется автоматизмом. Так как импульсы появляются в мышечных волокнах, то говорят о миогенной автоматии.
Существование миогенной автоматии позволяет возбуждаться и сокращаться сердечной мышце при перерезке всех идущих к ней внешних нервов и даже при полном извлечении сердца из тела. При создании необходимых условий, способность к сокращению, без действия внешних раздражителей, сохраняется в течение нескольких часов и даже суток. Ритмические сокращения зарегистрированы у человеческого эмбриона на ранних стадиях развития (18-20 день).
Но способностью к автоматии в сердце обладают не все мышечные волокна, а только атипическая мышечная ткань.
Природа автоматии до сих пор до конца не выяснена. У высших позвоночных возникновение импульсов связано с функцией атипических мышечных клеток - миоцитов -пейсмекеров , заложенных в узлах сердца.
Атипическая ткань в сердце млекопитающих животных локализуется в областях, гомологичных венозному синусу и атриовентрикулярной области холоднокровных.
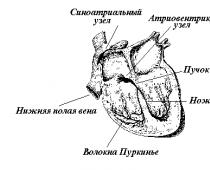
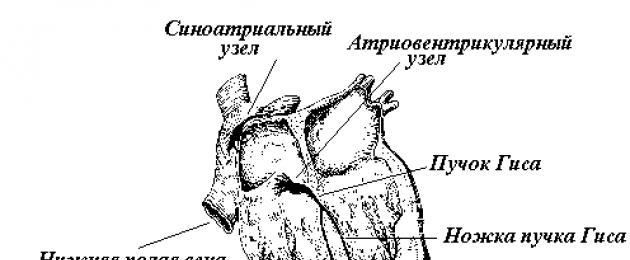
Первый узел проводящей системы расположен в месте впадения полых вен в правое предсердие. Имеет несколько названий: синуснопредсердный, синоатриальный, синусный, синусноаурикулярный, Кейс-Флека (Кис-Фляка, Кейт-Флака) . Он является главным центром автоматии сердца -пейсмекером (водителем ритма) первого порядка .
От данного узла возбуждение распространяется к рабочим клеткам миокарда, как диффузно, так и по специализированным пучкам или трактам (Торела, Венкебаха, Кента и т.д.).
В частности к левому предсердию возбуждение направляется по пучку Бахмана, а к предсердно-желудочковому узлу - по пучку Кис-Фляка.
Далее возбуждение достигает второго узла -предсердно-желудочкового (атриовентрикулярного, Ашоффа-Товара) . Он расположен в толще сердечной перегородки на границе предсердий и желудочков. Узел состоит из трех частей, обладающих собственной частотой возбуждения: 1 -верхней предсердной и 2-средней и 3 -нижней желудочковых. Этот узел является пейсмекером второго порядка . В норме возбуждение в данном узле никогда не генерируется, узел лишь проводит импульсы от синоатриального узла, причем в норме возбуждение проходит только в одном направлении. Ретроградное (обратное) проведение импульсов невозможно.
При прохождении возбуждения через предсердно-желудочковый узел импульсы задерживаются на 0,02-0,04 с. Это явление получило название атриовентрикулярной задержки . Ее функциональное значение состоит в том, что за время задержки успевает завершиться систола предсердий. За счет этого достигается координированная работа предсердий и желудочков.
В настоящее время предполагается, что причиной атриовентрикулярной задержки может быть: истончение пучков Кис-Флака при подходе к атриовентрикулярному узлу. Существует также предположение, что передача возбуждения на атриовентрикулярный узел осуществляется через химический синапс.
Третий уровень расположен в пучке Гиса и волокнах Пуркинье. Пучок Гиса берет начало от предсердно-желудочкового узла (длина 1-2 см) и образует две ножки, одна из которых идет к левому, другая - к правому желудочку. Эти ножки ветвятся на более тонкие проводящие пути, которые в свою очередь заканчиваются волокнами Пуркинье под эндокардом. Считается, что между этими волокнами и типичной мускулатурой имеются так называемые переходные клетки. Они непосредственно и осуществляют контакт с рабочими клетками миокарда и обеспечивают одновременную передачу возбуждения с проводящей системы сердца на рабочую мускулатуру.
Центры автоматии, расположенные в проводящей системе желудочков, носят название пейсмекеров третьего порядка . Они также, как и атриовентрикулярный узел, никогда в норме не вступают в работу, а предназначены лишь для проведения импульсов, идущих от синоатриального узла. Таким образом, возбуждение по ножкам пучка Гиса направляется к верхушке сердца и оттуда по разветвлениям ножек и волокнам Пуркинье возвращается к основанию сердца. В результате этого сокращение сердца в целом определяется в определенной последовательности: сначала сокращаются предсердия, затем верхушки желудочков и наконец их основания.
Итак, нижележащие водители ритма находятся в соподчиненном положении и в сердце существует так называемый градиент автоматии , который был открыт в опытах Станиуса (описаны в практических руководствах по физиологии), а сформулирован Гаскеллом.
Градиент автоматии выражается в убывающей способности к автоматии различных структур проводящей системы по мере их удаления от синусно-предсердного узла. В синусно-предсердном узле число разрядов составляет в среднем 60-80 имп/мин у взрослого человека, в предсердно-желудочковом - 40-50, в клетках пучка Гиса - 30-40, в волокнах Пуркинье - 20-30 имп/мин.
Таким образом, в сердце существует определенная иерархия центров автоматии, что позволило В. Гаскеллу сформулировать правило, согласно которому степень автоматии отдела тем выше, чем он ближе расположен к синусно-предсердному узлу.
В том случае, когда в пейсмекере первого порядка не возникает возбуждение или блокируется его передача, роль водителя ритма берет на себя пейсмекер второго порядка через 30-40 сек (асистолия) и желудочки начинают сокращаться в ритме атриовентрикулярного узла. При невозможности передачи возбуждения к желудочкам они начинают сокращаться в ритме пейсмекеров третьего порядка.
В норме частоту активности миокарда всего сердца в целом определяет синусно-предсердный узел и подчиняет себе все нижележащие центры автоматии, навязывая им свой ритм. Явление, при котором структуры с замедленным ритмом генерации потенциалов усваивают более частый ритм других участков проводящей системы, называют усвоением ритма. В случае, когда синоатриальный узел повреждается и при этом человеку оказывают своевременную квалифицированную медицинскую помощь (больному вживляют стимулятор, задающий самостоятельно ритм для работы сердца) можно сохранить жизнь пациенту.
При поперечной блокаде предсердия и желудочки сокращаются каждый в своем ритме. Нескоординированная работа водителей ритма ухудшает основную функцию сердца - нагнетательную. Повреждение водителей ритма ведет к полной остановке сердца.
Материалы для самостоятельной работы студентов
(Составители – ,)
СВОЙСТВА СЕРДЕЧНОЙ МЫШЦЫ
1. Механизмы электрогенеза миокардиальных клеток
Функциональным элементом сердца является мышечное волокно - цепочка из клеток миокарда, соединенных “конец в конец” и заключенных в общую саркоплазматическую оболочку.
Потенциал покоя (ПП) миокардиальных волокон формируется за счет высокой проницаемости поверхностной протоплазматической мембраны миокардиального волокна для катионов калия. Возникновение потенциалов действия (ПД) обусловлено открытием натриевых потенциалзависимых каналов поверхностной протоплазматической мембраны. Значительный вклад в генерацию ПД миокардиальных волокон вносят потенциалзависимые кальциевые каналы. Возбуждение распространяется по сердцу без декремента, механизм распространения - электрический.
Сердечная мышца неоднородна в своем клеточном составе. Различают типичные (сократительные) и атипичные волокна миокарда. Они различаются по строению, функции и электрической активности. DIV_ADBLOCK99">
Иначе выглядят мембранные потенциалы, регистрируемые в атипичных волокнах миокарда (АТМВ). В них нет устойчивого уровня поляризации. Электрическая активность АТМВ представляет собой непрерывные колебания мембранного потенциала. По достижении им определенной величины (примерно - 60 мВ) спонтанно начинает развиваться медленная диастолическая деполяризация . На уровне около -40 мВ процесс деполяризации ускоряется, кривая идет круче (рис. 1, А), что соответствует возникновению в АТМВ потенциала действия. Деполяризация сменяется реполяризацией, в конце которой вновь начинается медленная диастолическая деполяризация, лежащая в основе самопроизвольной ритмической активности сердца .
Свойство миокарда возбуждаться под влиянием ПД, спонтанно возникающих в нем самом, называется автоматизмом (автоматией). Электрические процессы, связанные с ним, происходят в АТМВ. Таким образом, атипичные миокардиальные волокна служат источником автоматизма сердечной мышцы.
2. Особенности распространения возбуждения по миокарду
Атипичная миокардиальная ткань сосредоточена в виде островков в различных отделах сердца, объединяющихся в проводящую систему сердца (рис. 2). Особенности проведения возбуждения по миокарду связаны со способом соединения между собой его отдельных волокон. Под электронным микроскопом между ними обнаружили особые вставочные диски. Они представляют собой дубликатуры мембран, в которых имеются щелевые контакты между соседними волокнами. Открытие вставочных дисков дало основание рассматривать миокард как функциональный синцитий, поскольку электрический импульс (ПД), возникший в одном волокне, распространяется на соседние посредством электрической передачи. Благодаря этому сердцу присущи свойства гомогенной возбудимой системы. Так, в реакциях на одиночные раздражения миокард подчиняется закону “все или ничего”. Электрическая связь существует не только между типичными мышечными волокнами сердца. Аналогично связаны атипичные миокардиальные волокна как между собой, так и с типичными мышечными волокнами. Поэтому возбуждение, ритмически возникающее в АТМВ, легко переходит на ТМВ и распространяется по всему сердцу.
Между скоплениями атипичной миокардиальной ткани установлена четкая субординация: ведущим является то скопление АТМВ, которое расположено в правом предсердии - между венозным синусом и ушком - синоатриальный узел . Когда он автоматически возбуждается, другие островки АТМВ выполняют только функцию проведения. Их собственный автоматизм угнетен. Поэтому синоатриальный узел называют пейсмекером - водителем ритма, так как ритм его автоматических возбуждений определяет частоту сердечных сокращений. Количественной мерой автоматизма различных АТМВ является частота спонтанных возбуждений. Чем она больше, тем выше уровень автоматизма. Основным водителем ритма служит группа из примерно 5000 атипичных волокон, занимающих около 3,5% объема этого узла. Отдельные волокна, входящие в группу, возбуждаются синхронно и синфазно, что обеспечивается электротоническим характером их взаимодействия. Передача возбуждения на другие группы АТМВ синоатриального узла, а также на ТМВ предсердий осуществляется посредством ПД. Благодаря тому, что водителем ритма является не одиночное АТМВ а группа волокон, сердце имеет высокую надежность в поддержании спонтанных колебаний.
С АТМВ синоатриального узла возбуждение переходит на ТМВ предсердий и проводится по ним со скоростью около 1 м/с. Уже через 40 мс от возникновения ПД в водителе ритма все участки предсердий находятся в возбужденном состоянии. Возбуждение распространяется по предсердиям широким фронтом, что обеспечивается наличием щелевых контактов не только между торцами отдельных миокардиальных волокон, имеющих продольное расположение, но и между их боковыми поверхностями. Однако плотность щелевых контактов в торцевых

Рис. 2. Схема строения проводящей системы сердца.
вставочных дисках выше, чем в боковых, поэтому по направлению к желудочкам возбуждение движется быстрее, чем поперек предсердий. Тем самым достигается сокращение всего миокарда предсердий с одномоментным выходом волны возбуждения на атриовентрикулярный узел проводящей системы сердца. Как известно, предсердия отделены от желудочков фиброзной тканью, которая не способна проводить возбуждение. Вместе с тем, в этой преграде есть узкая щель - шириной чуть более 1 мм и длиной 1,5-2 мм, в которой расположен атриовентрикулярный узел, проводящий возбуждение из предсердий в желудочки. В местах контакта с типичным миокардом предсердий АТМВ атриовентрикулярного узла очень тонки, вследствие чего им присуще значительное электрическое сопротивление саркоплазмы. В этом одна из причин резкого в 20-50 раз замедления распространения возбуждения в атриовентрикулярном узле по сравнению с предсердиями. Другая причина заключается в том, что АТМВ в верхней части узла имеют не продольное, а поперечное расположение. Следовательно, по направлению к желудочкам возбуждение передается через боковые, а не более эффективные торцевые вставочные диски.
Замедленное проведение возбуждения из предсердий в желудочки обеспечивает важную для нормальной работы сердца паузу между сокращениями. Ее называют атриовентрикулярной задержкой. Желудочки начинают сокращаться примерно через 0,1 с от начала сокращения предсердий. Задержка нужна для того, чтобы кровь, накопленная предсердиями в диастолу, полностью перешла в желудочки до того, как они начнут сокращаться, нагнетая ее в аорту.
Из атриовентрикулярного узла возбуждение поступает в пучок Гиса . Там скорость проведения возбуждения возрастает до 2-3 м/c. Увеличение скорости обусловлено утолщением АТМВ и повышением плотности щелевых контактов во вставочных дисках. Ближе к верхушке сердца от пучка Гиса отходят волокна Пуркинье . Эти атипичные миокардиальные волокна вступают в контакт с ТМВ желудочков. Волокна Пуркинье обладают наибольшим диаметром по сравнению с другими волокнами миокарда. Поэтому скорость проведения возбуждения здесь достигает 4-5 м/с. Возбуждение с отдельных волокон Пуркинье переходит на огромное число ТМВ практически одномоментно, благодаря чему разные участки желудочков сокращаются синфазно.
3. Электромеханическое сопряжение в миокарде
Сокращение миокардиальных волокон, как и волокон скелетных мышц, инициируется потенциалом действия. Тем не менее временные соотношения между параметрами потенциала действия и параметрами сокращения этих мышечных волокон различны. Длительность потенциала действия скелетных мышц составляет несколько миллисекунд, и сокращение их начинается после завершения развития потенциала действия. В миокарде потенциал действия и сокращение в значительной степени перекрываются во времени. Потенциал действия клеток миокарда заканчивается после начала фазы расслабления. Поскольку последующее сокращение может возникнуть только в результате очередного возбуждения, а это возбуждение в свою очередь возможно лишь по окончании периода абсолютной рефрактерности предшествующего потенциала действия, сердечная мышца в отличие от скелетной не может отвечать на частые раздражения суммацией одиночных сокращений, или тетанусом . Это свойство миокарда имеет большое значение для реализации нагнетательной функции сердца: с одной стороны - тетаническое сокращение, продолжающееся больше периода изгнания крови, препятствовало бы наполнению сердца, с другой - тетаническое сокращение сердца эквивалентно его остановке.
Невозможность сердечной мышцы давать тетанические сокращения заставило детально проанализировать вопрос о механизмах регуляции силы сердечных сокращений. Как было отмечено, сократимость сердца не может регулироваться путем суммации одиночных сокращений, сократимость миокарда в отличие от скелетных мышц, не может изменяться путем включения различного числа моторных единиц, так как миокард представляет собой функциональный синцитий, и в каждом его сокращении участвуют все волокна. Однако, эти несколько невыгодные с физиологической точки зрения особенности компенсируются тем, что в миокарде возможность регуляции сократимости обеспечивается путем направленного изменения процессов возбуждения и электромеханического сопряжения.
Как организован механизм электромеханического сопряжения в миокарде? У человека и у млекопитающих структуры, отвечающие за электромеханическое сопряжение в скелетных мышцах, в основном, имеются и в волокнах сердца. Для миокарда характерна система поперечных трубочек (Т-система); особенно хорошо она развита в желудочках, где эти трубочки образуют продольные ответвления. Напротив, система продольных трубочек, служащих внутриклеточным резервуаром Са2+, в мышце сердца выражена в меньшей степени, чем в скелетных мышцах. Как структурные, так и функциональные особенности миокарда свидетельствуют в пользу тесной взаимосвязи между внутриклеточными депо Са2+ и внеклеточной средой. Ключевым событием в сокращении служит вход в клетку Са2+во время потенциала действия. Значение входного кальциевого тока состоит не только в том, что он увеличивает длительность потенциала действия и, как следствие, рефракторного периода: перемещение Са2+ из наружной среды в клетку создает условия для регуляции силы сокращения, так как чем больше кальция оказывается вблизи актина и миозина, тем сильнее сокращается
Активация" href="/text/category/aktivatciya/" rel="bookmark">активацией сократительного аппарата. Начало сокращения связано с выходом кальция в зону актина и миозина из продольных трубочек в ходе деполяризации мембраны. Кальций, поступающий в кардиомиоцит через кальциевые каналы в фазу плато потенциала действия кардиомиоцита, пополняет запасы кальция в продольных трубочках.
На концентрацию кальция, активирующего контрактильный механизм, существенно влияет его количество в продольных трубочках, при этом показано, что значительная часть входящего в клетку Са2+ пополняет его запасы, обеспечивая достаточную эффективность очередных сокращений.
Таким образом, потенциал действия влияет на сократимость по меньшей мере двумя путями. 1. Он играет роль пускового механизма, вызывающего сокращение путем высвобождения Са2+ преимущественно из внутриклеточных депо. 2. Он пополняет запасы Са2+, создавая благоприятные условия для последующих сокращений.
Как выяснилось, целый ряд агентов оказывает значимое влияние на сокращение миокарда, изменяя длительность потенциала действия, что отражается на поступлении Са2+ внутрь миокардиоцитов. Например, ацетилхолин, выделяющий в окончаниях блуждающего нерва, уменьшая продолжительность потенциалов действия предсердной мышцы, параллельно усиливая проницаемость поверхностной мембраны клеток синоатриального узла для калия, вызывая тем самым их гиперполяризацию и уменьшение входного тока Са2+, вызывает уменьшение частоты и силы сердечных сокращений (отрицательный хронотропный, инотропный, дромотропный эффекты - см. ниже.). Напротив, норадреналин, выделяющийся в окончаниях симпатических нервных волокон, усиливая проницаемость для Са2+ , вызывает повышение частоты и увеличение силы сердечных сокращений (положительный хронотропный, инотропный, дромотропный эффекты - см. ниже).
Так называемый феномен лестницы (нарастание силы сокращений при их возобновлении после временной остановки) также связан с наращиванием внутриклеточной фракции Са2+. Сила сокращения сердца быстро изменяется при изменении содержания Са2+ во внеклеточной жидкости. Удаление Са2+ из внешней среды приводит к полному электромеханическому разобщению. Ряд веществ, блокирующих вход Са2+ во время потенциала действия, оказывает такой же эффект, как и удаление Са2+ из внешней среды. К таким веществам относятся двухвалентные катионы (Ni 2+ , Со2+, Mn 2+ ), а также некоторые органические соединения - антагонисты кальция (верапамил, нифедипин). При повышении внеклеточного содержания Са2+ или при действии фармакологических препаратов, увеличивающих вход Са2+ во время развития потенциала действия, сократимость сердца увеличивается. Механизм действия сердечных гликозидов (дигоксин, строфантин) частично связан именно с увеличением внутриклеточной фракции Са2+.
4. Биофизические основы электрокардиографии
Особенности распространения возбуждения по сердцу отображаются в электрокардиограмме (ЭКГ ), которая имеет характерную форму (рис. 4). Элементарной моделью генератора ЭКГ является электрический диполь. При распространении возбуждения по миокарду формируется множество диполей, которые закономерно изменяют свои количественные характеристики и направление. В каждый момент времени возникают новые диполи, исчезают прежние. В результате на поверхности сердечной мышцы создается сложная мозаика распределения электрических потенциалов. Результирующий дипольный момент миокарда, равный векторной сумме отдельных диполей, получил название интегрального электрического вектора сердца (ИЭВС). Ритмичный характер автоматизма водителя ритма, а также передача возбуждения посредством электрических синапсов обусловливают синфазность возбудительного процесса в миокардиальных волокнах. Поэтому ИЭВС имеет сравнительно большую амплитуду прежде всего при деполяризации желудочков, чем создается высокий уровень биопотенциалов, отражающих сердечную деятельность даже на поверхности тела. Ежемоментно амплитуда и направление ИЭВС различны. Измеряя их, врач получает сведения о движении волны возбуждения по сердцу, что позволяет ему оценить свойства миокарда и в случае нарушений сердечной деятельности понять их природу.
https://pandia.ru/text/80/111/images/image005_20.jpg" width="306 height=259" height="259">
Рис. 5. Соотношение векторэлектрокардиограммы (А) и электрокардиограммы (Б).
В данном случае фигуры Лиссажу представляют собой траектории движения ИЭВС, описываемые его концом на плоскости, перпендикулярной направлению распространения возбуждения по миокарду. Такое исследование электрической активности сердца называется векторэлектрокардиоскопией (ВЭКС ). На векторэлектрокардиограмме выделяют обычно три эллипсоподобные фигуры. Самая мелкая из них отображает деполяризацию предсердий, самая крупная - деполяризацию желудочкой, средняя - их реполяризацию.
Амплитуду вектора оценивают посредством измерения его проекций на координатные оси. Любое из отведений ЭКГ есть не что иное как проекция интегрального электрического вектора сердца на соответствующую координатную ось.
DIV_ADBLOCK101">
Важным параметром ЭКГ служат временные интервалы. По ним оценивают скорость распространения возбуждения в каждом из отделов проводящей системы сердца. Изменения скорости проведения наблюдаются при повреждениях сердечной мышцы. Даже мелкий очаг поражения миокарда (диаметром 5-10 мкм) вызывает задержку в распространении возбуждения на 0,1 мс.
В стандартных отведениях зубец Р имеет амплитуду не более 0,25 мВ и длительность 0,07-0,10 с. Интервал PQ, отображающий атрио-вентрикулярную задержку, составляет 0,12-0,21 с при частоте сердечных сокращений порядка 70 /мин. Комплекс QRS наблюдается в течение всего времени, пока возбуждение распространяется по желудочкам - от 0,06 до 0,09 с. Зубец Q в трети наблюдений отсутствует в нормальной ЭКГ, а когда обнаруживается, не превышает 0,25 мВ. Зубец R обладает максимальной амплитудой среди других элементов ЭКГ. Она составляет 0,6-1,6 мВ. Зубец S -часто отсутствует, но иногда достигает 0,6 мВ. Он появляется в тот момент, когда деполяризация охватывает участки желудочков, прилежащие к предсердиям. Основание желудочков возбуждается в последнюю очередь. Сегмент S-T при пульсе 65-70 /мин составляет 0,12 с. Длительность зубца Т немного больше - от 0,12 до 0,16 с. Его амплитуда находится в пределах 0,25-0,6 мВ.
Зубец Р возникает на ЭКГ примерно за 0,02 с до начала сокращения предсердий, а комплекс QRS - за 0,04 с до начала сокращения желудочков. Следовательно, электрические проявления возбуждения предшествуют меха-ническим.
Имея ряд ЭКГ, по крайней мере, две, снятые в 1 и 3 отведениях, можно синтезировать ИЭВС. В медицинской литературе его называют электрической осью сердца - отрезок прямой, соединяющий два сечения миокарда, облада-ющих в данный момент наибольшей разностью потенциалов. Направление электрической оси сердца в ходе распространения возбуждения по миокарду постоянно изменяется. Принято определять среднюю электрическую ось сердца. Так называют вектор, который можно построить в промежутке между началом и окончанием деполяризации миокарда желудочков. По расположению средняя электрическая ось близка анатомической оси сердца. Построение средней электрической оси дает представление о положении сердца в грудной полости. Отклонения оси вправо или влево служат признаками изменений миокарда соответствующего желудочка.
Автоматия - способность сердечной мышцы к ритмическому сокращению без всяких внешних воздействий под влиянием импульсов, возникающих в самом сердце. Благодаря автоматии автономное (извлеченное из организма) сердце способно некоторое время самостоятельно сокращаться. Импульсы в сердечной мышце возникают благодаря деятельности атипических мышечных волокон, заложенных в некоторых участках миокарда - внутри них спонтанно генерируются электрические импульсы определенной частоты, распространяющиеся затем по всему миокарду. Первый такой участок находится в области устьев полых вен и называется синусовым , или синоатриальным, узлом. Он производит импульсы с частотой 60-80 раз в минуту и является главным центром автоматии сердца. Второй участок находится в толще перегородки между предсердиями и желудочками и называется предсердно-желудочковым, или атриовентрикулярным , узлом. Третий участок - пучок Гиса - атипические волокна, лежащие в межжелудочковой перегородке. От пучка Гиса отходят тонкие волокна атипической ткани - волокна Пуркинье, разветвляющиеся в миокарде желудочков. Все участки атипической ткани способны самостоятельно генерировать импульсы; в синусовом узле их частота самая высокая, его называют водителем ритма первого порядка, другие центры автоматии подчиняются этому ритму. Совокупность всех центров автоматии составляют проводящую систему сердца, благодаря которой волна возбуждения, возникшая в синусном узле, последовательно распространяется по всему миокарду и обеспечивает последовательное сокращение отделов сердца.
Возбудимость сердечной мышцы проявляется в способности сердца приходить в состояние возбуждения под действием различных раздражителей (химических, механических, электрических и др.). Потенциал действия, возникающий в одной клетке, передается другим клеткам, что приводит к распространению возбуждения по всему сердцу.
Сократимость - способность полости сердца сокращаться, обусловленная свойством клеток миокарда отвечать на возбуждение сокращением. Это свойство сердечной мышцы позволяет сердцу выполнять механическую работу по перекачиванию крови по сосудам: при сокращении полости сердца давление крови в сердечных камерах возрастает, и кровь под давлением поступает в артерии. Работа сердечной мышцы подчиняется закону «все или ничего»: если на сердечную мышцу оказывать раздражающее действие различной силы, мышца каждый раз отвечает максимальным сокращением. Если сила раздражителя не достигает порогового значения, то сердечная мышца не отвечает сокращением.
В работе сердца как насоса выделяют три фазы, сокращение предсердий, сокращение желудочков и пауза, когда желудочки и предсердия одновременно расслаблены. Сокращение сердца называется систолой , расслабление - диастолой. Во время систолы предсердий кровь выталкивается в желудочки, так как обратный кровоток в вены невозможен из-за захлопывания клапанов, во время систолы желудочков кровь устремляется в большой и малый круги кровообращения (обратному току в предсердия препятствуют митральный и трехстворчатый клапаны, расположенные между предсердиями и желудочками), а за время диастолы камеры сердца находятся в расслабленном состоянии и вновь заполняются кровью. За одну минуту сердце взрослого здорового человека сокращается примерно 60-70 раз. Ритмичное чередование сокращения и расслабления каждого из отделов сердца обеспечивает неутомляемость сердечной мышцы.
Иннервация сердца очень сложна. Она осуществляется вегетативной нервной системой - блуждающим и симпатическими нервами, в составе которых имеются как чувствительные, так и двигательные волокна. В стенке самого сердца находятся нервные сплетения, состоящие из нервных узлов и нервных волокон. Двигательные нервы сердца осуществляют четыре основные функции: замедление, ускорение, ослабление и усиление деятельности сердца. Эти нервы относятся к вегетативной нервной системе. Таким образом, сердечная мышца, обладая способностью к самостоятельным сокращениям, подчиняется также «командам сверху» - регулирующему воздействию нервной системы, обеспечивающему оптимальную адаптацию сердечной деятельности потребностям организма в конкретной ситуации.
Сосудистая система. Кровеносные сосуды представляют собой систему полых эластичных трубок различного строения, диаметра и механических свойств, по которым протекает кровь. Сосуды подразделяются на артерии, вены и капилляры.
Артерии имеют толстые упругие стенки, состоящие из грех слоев. Наружный слой представляет собой соединительнотканную оболочку, средний слой состоит из гладкой мышечной ткани и содержит соединительнотканные эластические волокна, внутренний слой образован эндотелием, под которым расположена внутренняя эластическая мембрана. Эластические элементы артериальной стенки образуют единый каркас, работающий как пружина и обусловливающий эластичность артерий.
Разветвляясь, артерии переходят в артериолы , которые отличаются от артерий наличием только одного слоя мышечных клеток и могут регулировать скорость кровотока за счет сужения или расширения просвета. Артериола переходит в прекапилляр, в котором мышечные клетки разрознены и не составляют сплошного слоя. От него отходят многочисленные капилляры - самые мелкие кровеносные сосуды, которые соединяют артериолы с венулами (мелкими разветвлениями вен). Благодаря очень тонкой стенке капилляров в них происходит обмен различными веществами между кровью и клетками тканей. В зависимости от потребности в кислороде и других питательных веществах разные ткани имеют разное количество капилляров. Капилляры могут находиться в активном (открытом) и пассивном (закрытом) состоянии. При активизации обменных процессов или потребности в усиленной теплоотдаче объем крови, проходящей через орган, может увеличиваться за счет активизации дополнительного числа капилляров. В покое и при уменьшении теплоотдачи значительное количество капилляров переходит в пассивное состояние, уменьшая таким образом объем кровотока. Состояние капиллярной сети регулируется вегетативной нервной системой в зависимости от потребностей организма.
Сливаясь, капилляры переходят в посткапилляры , которые но строению аналогичны прекапилляру. Посткапилляры сливаются в венулы с просветом 40-50 мкм. Венулы объединяются в более крупные сосуды, несущие кровь к сердцу, - вены. Они, так же как и артерии, имеют стенки, состоящие из трех слоев, по содержат меньше эластических и мышечных волокон, поэтому менее упруги, их просвет поддерживается током крови. Вены имеют клапаны (полулунные складки внутренней оболочки), которые открываются по току крови, что способствует движению крови в одном направлении. Схематически строение кровеносных сосудов представлено на рис. 4.6.
Рис. 4.6.
Человек и все позвоночные животные имеют замкнутую кровеносную систему. Кровеносные сосуды сердечно-сосудистой системы образуют две основные подсистемы: большой и малый круги кровообращения (рис. 4.7).
Сосуды большого круга кровообращения соединяют сердце со всеми другими частями тела. Большой круг кровообращения начинается в левом желудочке, откуда выходит аорта, а заканчивается в правом предсердии, куда впадают полые вены. Как часть большого круга кровообращения выделяют третий (сердечный) круг, снабжающий кровью само сердце. Он состоит из двух венечных, или коронарных, артерий, отходящих от аорты, и впадает в правое предсердие через венечную пазуху.
Сосуды малого круга кровообращения переносят кровь от сердца к легким и обратно. Малый круг кровообращения начинается правым желудочком, из которого выходит легочный ствол, а заканчивается левым предсердием, в которое впадают легочные вены.

Рис. 4.7.
1 - сердце; 2 - малый (легочный) круг кровообращения; 3 - большой круг кровообращения
Может выполнять свои многочисленные функции, только находясь в постоянном движении. Обеспечение движения крови является главной функцией сердца и сосудов, формирующих кровеносную систему. Сердечно-сосудистая система совместно с кровью участвует также в транспорте веществ, терморегуляции, реализации иммунных реакций и гуморальной регуляции функций организма. Движущая сила кровотока создастся за счет , которое выполняет функцию насоса.
Способность сердца сокращаться в течение всей жизни без остановки обусловлена рядом специфических физических и физиологических свойств сердечной мышцы. Сердечная мышца уникальным образом сочетает в себе качества скелетной и гладкой мускулатуры. Так же как и скелетные мышцы, миокард способен интенсивно работать и быстро сокращаться. Так же как и гладкие мышцы, он практически неутомим и не зависит от волевого усилия человека.
Физические свойства
Растяжимость — способность увеличивать длину без нарушения структуры под влиянием растягивающей силы. Такой силой является кровь, наполняющая полости сердца во время диастолы. От степени растяжения мышечных волокон сердца в диастолу зависит сила их сокращения в систолу.
Эластичность - способность восстанавливать исходное положение после прекращения действия деформирующей силы. Эластичность сердечной мышцы является полной, т.е. она полностью восстанавливает исходные показатели.
Способность развивать силу в процессе сокращения мышцы.
Физиологические свойства
Сокращения сердца происходят вследствие периодически возникающих процессов возбуждения в сердечной мышце, которая обладает рядом физиологических свойств: , .
Способность сердца ритмически сокращаться под влиянием импульсов, возникающих в нем самом, носит название автоматизм.
В сердце различают сократительную мускулатуру, представленную поперечно-полосатой мышцей, и атипическую, или специальную ткань, в которой возникает и проводится возбуждение. Атипическая мышечная ткань содержит малое количество миофибрилл, много саркоплазмы и не способна к сокращению. Она представлена скоплениями в определенных участках миокарда, которые образуют , состоящую из синоатриального узла, располагающегося на задней стенке правого предсердия у места впадения полых вен; атриовентрикулярного, или предсердно-желудочкового узла, находящегося в правом предсердии вблизи перегородки между предсердиями и желудочками; предсердно-желудочкового пучка (пучка Гиса), отходящего от атриовентрикулярного узла одним стволом. Пучок Гиса, пройдя через перегородку между предсердиями и желудочками, разветвляется на две ножки, идущие к правому и левому желудочкам. Заканчивается пучок Гиса в толще мышц волокнами Пуркинье.
Синоатриальныи узел является водителем ритма первого порядка. В нем возникают импульсы, которые определяют частоту сокращений сердца . Он генерирует импульсы со средней частотой 70-80 импульсов в 1 мин.
Атриовентрикулярный узел - водитель ритма второго порядка.
Пучок Гиса - водитель ритма третьего порядка.
Волокна Пуркинье — водители ритма четвертого порядка. Частота возбуждения, возникающая в клетках волокон Пуркинье, очень низкая.
В норме атриовентрикулярный узел и пучок Гиса являются только передатчиками возбуждений из ведущего узла к сердечной мышце.
Однако и они обладают автоматизмом, только в меньшей степени, и этот автоматизм проявляется лишь при патологии.
В области синоатриального узла обнаружено значительное число нервных клеток, нервных волокон и их окончаний, которые образуют здесь нервную сеть. К узлам атипической ткани подходят нервные волокна от блуждающих и симпатических нервов.
Первыми сокращаются мышцы предсердий, затем слой мышц желудочков, обеспечивая тем самым движение крови из полостей желудочков в аорту и легочный ствол.