Мышечная ткань выполняет следующие функции:
Обеспечение двигательной активности – целенаправленное поведение есть наиболее эффективная форма приспособления.
Обеспечение особых, присущих только человеку функций – прежде всего это коммуникативная функция , выражающаяся в виде устной и письменной речи.
Выполнение дыхательной функции – экскурсия грудной клетки и диафрагмы.
Участие в процессах теплообразования – терморегуляторный тонус, мышечная дрожь.
Специфическим свойством всех типов мышц является сократимость – способность сокращаться, то есть укорачиваться или развивать напряжение. Для реализации этой способности мышца использует два своих дополнительных свойства – возбудимость и проводимость .
Скелетные мышцы называют также произвольными , так как их сокращением можно управлять по собственному желанию. Они полностью лишены автоматизма и не способны работать без управляющей импульсации из ЦНС. Гладкие мышцы по собственному желанию не сокращаются, поэтому их называют также непроизвольными .
Морфофункциональная характеристика скелетной мышцы . Скелетная мышца состоит из многоядерных мышечных волокон. Толщина волокна составляет от 10 до 100 мкм. Длина волокон колеблется от нескольких мм до нескольких сантиметров.
Количество мышечных волокон становится постоянным на 4-5 месяце постнатального развития. В последующем увеличивается лишь диаметр и длина волокон (например, под действием тренировок – функциональная гипертрофия).
Мышечное волокно покрыто сарколеммой. В саркоплазме мышечного волокна имеются следующие внутриклеточные элементы: ядра, митохондрии, белки, капельки жира, гранулы гликогена, фосфатсодержащие вещества, различные малые молекулы и электролиты. От поверхности сарколеммы внутрь волокна отходят Т-трубочки, которые обеспечивают её взаимодействие с саркопламатическим ретикулумом. Саркоплазматический ретикулум представляет собой систему связанных между собой цистерн и отходящих от них в продольном направлении канальцев , расположенных между миофибриллами. Крайние цистерны ретикулума связаны с Т-трубочками. В цистернах содержатся ионы кальция, необходимые для осуществления процесса сокращения. Внутри мышечного волокна тянется масса нитей – миофибрилл, которые являются частью механизма процесса сокращения. Каждая миофибрилла состоит их протофибрилл, которые расположены параллельно друг другу и имеют белковую природу.
Различают два вида внутримышечных нитей: тонкие актиновые и толстые миозиновые . Актиновые нити состоят из двух субъединиц - скрученных в виде спирали волокон, каждое их которых образовано соединенными молекулами глобулярного белка актина. Кроме актина в состав тонких нитей входят регуляторные белки тропомиозин и тропонин . Эти белки в невозбужденной мышце препятствуют связи актина и миозина, поэтому мышца в покое находится в расслабленном состоянии.
Рис.1. Схема пространственной организации сократительных и регуляторных белков в исчерченной мышце.
Каждая миозиновая нить окружена шестью актиновыми нитями. Эти нити образуют своего рода цилиндр, внутри которого располагается миозиновая нить. Поперечные мостики миозиновой нити направлены в разные стороны, поэтому они взаимодействуют со всеми актиновыми протофибриллами. В свою очередь, каждая нить актина контактирует с тремя миозиновыми филаментами.
Гладкие мышцы содержат актиновые и миозиновые нити, имеющие химические характеристики, подобные актиновым и миозиновым нитям скелетных мышц. Но в гладких мышцах нет тропонинового комплекса, необходимого для запуска сокращения скелетной мышцы, следовательно, механизм инициации сокращения в них другой. Этот механизм подробно обсуждается далее в нашей статье.
Химические исследования показали, что актиновые и миозиновые нити , извлеченные из гладких мышц, взаимодействуют друг с другом во многом так же, как и в скелетной мышце. Более того, процесс сокращения активируется ионами кальция, а энергия для сокращения обеспечивается разрушением АТФ до АДФ.
Существуют, однако, значительные различия в морфологической организации гладких и скелетных мышц , а также в сопряжении возбуждения и сокращения, механизме запуска ионами кальция сократительного процесса, длительности сокращения и количестве энергии, необходимой для сокращения.
Морфологическая основа сокращения гладких мышц
Гладкие мышцы не имеют такой упорядоченной организации актиновых и миозиновых нитей, которая обнаруживается в скелетных мышцах, придавая им «полосатость». С помощью техники электронной микрофотографии выявляется гистологическая организация. Видно большое число актиновых нитей, прикрепленных к так называемым плотным тельцам. Некоторые из этих телец прикрепляются к клеточной мембране, другие распределяются внутри клетки. Некоторые из мембранных плотных телец соседних клеток связываются вместе мостиками из внутриклеточных белков. Через эти мостики в основном передается сила сокращения от одной клетки к другой.
В мышечном волокне среди актиновых нитей разбросаны миозиновые нити. Их диаметр более чем в 2 раза превышает диаметр актиновых нитей. На электронных микрофотографиях актиновых нитей обычно обнаруживают в 5-10 раз больше, чем миозиновых.
На рисунке представлена предполагаемая структура отдельной сократительной единицы внутри гладкомышечной клетки , где видно большое число актиновых нитей, исходящих от двух плотных телец; концы этих нитей перекрывают миозиновую нить, расположенную посередине между плотными тельцами. Эта сократительная единица похожа на сократительную единицу скелетной мышцы, но без специфической регулярности ее структуры. В сущности, плотные тельца гладкой мышцы играют ту же роль, что и Z-диски в скелетной мышце.
Существует и другое различие. Большинство миозиновых нитей имеют поперечные мостики с так называемой боковой полярностью. Мостики организованы следующим образом: на одной стороне они шарнирно фиксируются в одном направлении, а на другой - в противоположном направлении. Это позволяет миозину тянуть актиновую нить с одной стороны в одном направлении, одновременно продвигая с другой стороны другую актиновую нить в противоположном направлении. Такая организация позволяет гладкомышечным клеткам сокращаться с укорочением до 80% их длины вместо укорочения менее чем на 30%, характерного для скелетной мышцы.
Большинство скелетных мышц сокращаются и расслабляются быстро, но сокращения гладких мышц в основном являются длительными тоническими сокращениями, которые иногда продолжаются в течение нескольких часов или даже дней. Следовательно, можно ожидать, что морфологические и химические особенности гладких мышц должны отличаться от соответствующих характеристик скелетных мышц. Далее обсуждаются некоторые из этих отличий.
Медленная циклическая активность миозиновых поперечных мостиков . В гладкой мышце по сравнению соскелетной гораздо меньше скорость циклической активности миозиновых поперечных мостиков, т.е. скорость их прикрепления к актину, отсоединение от актина и повторное прикрепление для осуществления следующего цикла. Фактически частота циклов составляет лишь от 1/10 до 1/300 этого показателя в скелетной мышце. Однако, как считают, в гладкой мышце значительно больше относительное количество времени, в течение которого поперечные мостики остаются прикрепленными к актиновым нитям, что является главным фактором, определяющим силу сокращения. Возможной причиной медленного циклирования является гораздо меньшая по сравнению со скелетной мышцей АТФ-азная активность головок поперечных мостиков, в связи с чем скорость разрушения АТФ - источника энергии для движения головок поперечных мостиков - значительно снижена с соответствующим замедлением скорости их циклов.
Гладкие мышцы образуют стенки (мышечный слой) внутренних органов и кровеносных сосудов.
Микроскопическое строение гладких и поперечнополосатых мышц различно.
Физиологические свойства гладких мышц в связи с особенностями их строения и уровня обменных процессов значительно отличаются от физиологических свойств поперечнополосатых мышц.
Гладкие мышцы менее возбудимы, чем поперечнополосатые. Возбуждение в гладких мышцах может передаваться с одного волокна на другое, в отличие от нервных волокон и волокон поперечнополосатых мышц. Возбуждение по гладким мышцам распространяется с небольшой скоростью - 2-15 см/с.
Сокращение гладкой мускулатуры происходит более медленно и длительно. Так, сокращение гладкой мускулатуры кишечника кролика может продолжаться 5 с, еще более медленно протекает расслабление.
Вследствие продолжительности сократительного акта гладкая мышца даже под влиянием редких раздражителей может переходить в состояние длительного сокращения, которое напоминает тетанус скелетных мышц. Характерными для гладких мышц являются также длительные тонические сокращения.
Рефракторный период в гладких мышцах более продолжителен, чем в скелетных.
Важным свойством гладкой мышцы является ее большая пластичность, т.е. способность сохранять созданную растяжением длину без изменения напряжения. Данное свойство имеет существенное значение, так как некоторые органы брюшной полости (матка, мочевой пузырь, желчный пузырь) иногда значительно растягиваются.
Характерной особенностью гладких мышц является их способность к автоматии, которая обеспечивается нервными элементами, заложенными в стенках гладкомышечных органов.
Адекватным раздражителем для гладких мышц является их быстрое и сильное растяжение, что имеет большое значение для функционирования многих гладкомышечных органов (мочеточник, кишечник и другие полые органы).
Особенностью гладких мышц является также их высокая чувствительность к некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин и другие).
Гладкие мышцы иннервируются симпатическими и парасимпатическими вегетативными нервами, которые, как правило, оказывают противоположное влияние на их функциональное состояние.
Т.О. Продолжительность одиночного мышечного сокращения - 0,1с. Приблизительно фаза укорочения и расслабления для скелетной мышцы одинаково - 0,05с. ЛП длинее, чем ПД.
В гладких мышцах продолжительность от нескольких секунд до нескольких минут. Продолжительность фазы расслабления более продолжительнее. ЛП короче, чем ПД.
РАЗДЕЛ: ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
ЗАНЯТИЕ №1
ТЕМА: БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ В ОРГАНИЗМЕ.
ПОТЕНЦИАЛ ПОКОЯ, ПОТЕНЦИАЛ ДЕЙСТВИЯ.
ЗАКОНЫ ВОЗБУЖДЕНИЯ
Продолжительность занятия – 2 часa.
План и организация занятия.
1. Подготовительный этап занятия:
а) организационные мероприятия - 5 мин.
б) проверка и коррекция исходного уровня знаний, посредством разбора материала в устной форме или с использованием учебника - 20 мин.
2. Основной этап занятия:
а) выполнение практических работ - 45 мин.
б) запись протокола исследования - 15 мин.
в) анализ результатов исследования - 10 мин.
3. Конечный этап занятия:
а) контроль конечного уровня усвоения учебного материала тестовым контролем или решением ситуационных задач - 20 мин.
3. Учебные цели занятия.
ЗНАТЬ:
1. Понятия возбудимости и раздражимости.
2. Роль, значение и функции плазматической мембраны клеток.
3. Учение о потенциал зависимых натриевых, калиевых, хлорных, кальциевых каналах.
4. Учение о неравномерном распределении ионов в возбудимых тканях, о трансмембранном электрохимическом градиенте и равновесном потенциале.
5. Мембранно-ионные механизмы происхождения, физические характеристики и физиологическую роль потенциала покоя.
6. Механизм потенциала действия, как проявление распространяющегося возбуждения. Динамику ионных токов при возбуждении.
7. Ионную природу локального ответа и физиологические характеристики, отличающие локальный ответ от распространяющегося возбуждения.
8. Изменение возбудимости в различные фазы генерации потенциала действия. Объяснение лабильности.
9. Закон электротонического потенциала: процессы, происходящие под катодом и анодом, при внеклеточном действии постоянного тока на возбудимые ткани.
10. Законы «силы», «все или ничего», «силы-времени». Аккомодация каналов мембран клеток.
11. Понятие реобазы, хронаксии.
12. История учения об электрических явлениях в возбудимых тканях.
УМЕТЬ:
1. Рисовать схемы развития потенциала покоя и потенциала действия.
2. Рисовать кривые потенциала действия и изменения возбудимости клетки во время его генерации.
4. Рисовать схему эквивалентной электрической модели плазматической мембраны.
5. Приготовлять нервно-мышечный препарат лягушки.
6. Работать с измерительными приборами.
4. Методика проведения занятия:
1.Подготовительный этап занятия.
В начале занятия следует сформулировать его цель и задачи, что студенты должны знать и уметь по окончанию занятия. В соответствии с этим, необходимо объяснить студентам, что знание материала этой темы потребуется для понимания значения роли плазматической мембраны в механизмах функционирования всех клеток организма, а особенно они важны при изучении физиологических свойств и особенностей нервной, мышечной и секреторной тканей. Знание особенностей строения и видов транспорта через плазматическую мембрану позволит студентам объяснить происхождение и поддержание на должном уровне основных констант клеток, механизмов действия гормонов, медиаторов и лекарственных веществ, развитие процессов возбуждения и торможения в клетках организма и выполнение других специфических функций. Все полученные знания будут необходимы при изучении других разделов физиологии, при обучении на последующих теоретических и клинических кафедрах. Следует обратить внимание студентов на то, что в настоящее время основные исследования в мире в области физиологии проводятся на клеточном, мембранном или молекулярном уровне, что без знания этих разделов невозможно объяснить и понять причины различных заболеваний и проводить необходимую терапию.
Основную часть подготовительного этапа занятия необходимо посвятить контролю исходного уровня знаний студентов путем устного или тестового опроса.
2. Основной этап занятия.
Этот этап занятия следует посвятить разбору и коррекции исходного уровня знания студентов, с учетом проведенного контроля. С этой целью рекомендуется провести устный разбор материала по основным вопросам занятия и предложить студентам написать и нарисовать основные формулы, графики и схемы. В процессе разбора учебного материала необходимо выяснить все вопросы занятия, записать основные понятия и формулировки, зарисовать в отчеты схемы, графики и формулы. При этом студенты могут пользоваться любой учебной литературой: учебниками, справочниками, атласами, электронным учебником и другими источниками информации.
Практическая часть : Проведение лабораторных работ в соответствии с рабочей программой.
3. Заключительный этап занятия.
На этом этапе занятия проводится контроль конечного уровня знания студентов, для чего рекомендуется использовать либо тестовый контроль знаний, либо решение ситуационных задач.
В заключение занятия преподаватель проверяет и подписывает протоколы студентов по выполнению лабораторных работ, задает задание для самостоятельной подготовки к следующему занятию.
Лабораторные работы.
1. Приготовление нервно-мышечного препарата лягушки.
Для изучения физиологических свойств мышц и нервов часто используют нервно-мышечный препарат, приготовленный из задних лапок лягушки. Классическим нервно-мышечным препаратом считают икроножную мышцу и седалищный нерв, который ее иннервирует.
Ход работы . Отпрепаровав нерв до коленного сустава, перерезают конечность выше и ниже коленного сустава и получают нервно-мышечный препарат. Для приготовления препарата изолированной мышцы от нервно-мышечного препарата отсекают нерв.
2. Проводимость нерва и её нарушение.
Одним из основных физиологических свойств возбудимых тканей является возбудимость, которая у различных тканей различна. Для характеристики уровня возбудимости служит порог раздражения, т.е. минимальная сила раздражителя, при действии которой возникает ответная реакция. В экспериментальных условиях для определения возбудимости мышцы применяют прямой метод ее раздражения, т.е. раздражение, наносимое непосредственно на мышцу. Возбудимость нерва исследуют раздражением нерва, иннервирующего данную мышцу, т.е. методом непрямого раздражения мышцы.
Ход работы . Готовят нервно-мышечный препарат. Подавая на нерв одиночные стимулы с постоянной длительностью, например 0,5 мс, постепенно увеличивают амплитуду и находят ту минимальную силу раздражителя, которая вызывает едва заметное сокращение мышцы - это и будет порог раздражения для нерва.
Для определения порога раздражения мышцы на нее наносят прямое раздражение через проводящую цепь миографа, соединенного со стимулятором. Порог раздражения находят так же, как при непрямом раздражении.
Рекомендации к оформлению работы. Зарисуйте схему установки для прямого и непрямого раздражения мышцы, запишите результаты опыта и дайте сравнительную оценку возбудимости нерва и мышцы. Сделать выводы о различии величин возбудимости нерва и мышца.
3. Опыты Гальвани.
Первый опыт Гальвани.
Ход работы . Готовят препарат двух задних лапок лягушки и подвешивают его на штативе. Берут биметаллический пинцет, одна бранша которого сделана из меди, а другая - из железа. Медную браншу подводят под нервные сплетения, а другую прикладывают к мышцам лапки. Наблюдают сокращение мышц лапок.
Второй опыт Гальвани (сокращение без металла).
Вторым опытом Гальвани впервые было доказано существование в тканях «животного электричества», которое возникает между поврежденной и неповрежденной поверхностями мышцы. Если эти два участка соединить нервом нервно-мышечного препарата, то возникает ток покоя, который раздражает нерв и вызывает сокращение мышцы.
Ход работы . Набрасывают седалищный нерв таким образом, чтобы он одновременно коснулся поврежденной и неповрежденной поверхности мышц бедра. При этом происходит сокращение мышц голени.
4. Опыт Маттеучи.
Опыт Маттеучи.
Раздражение нерва токами действия скелетной мышцы (вторичный тетанус). Маттеучи в 1840 г. показал, что сокращение мышцы нервно-мышечного препарата может наступить, если нерв этого препарата набросить на сокращающиеся мышцы другого нервно-мышечного препарата. На основании этого было сделано заключение, что в мышце при ее возбуждении возникают токи, которые могут стать раздражителем для другого нервно-мышечного препарата. Эти токи были названы, токами действия.
Ход работы . Нерв одного нервно-мышечного препарата (с кусочком позвоночника) с помощью стеклянного крючка помещают на электроды, которые соединены со стимулятором. На мышцы этого препарата в продольном направлении набрасывают нерв второго нервно-мышечного препарата. Нерв первого нервно-мышечного препарата подвергают ритмическому раздражению. Наблюдают тетаническое сокращение обеих лапок.
5. Зависимость силы ответа от силы раздражителя.
Скелетная мышца на раздражители пороговой силы отвечает минимальным пороговым сокращением. Если силу раздражителя постепенно увеличивать, то амплитуда сокращений скелетной мышцы также будет постепенно возрастать от пороговых до субмаксимальных и максимальных сокращений, после чего увеличение силы раздражителя не вызывает дальнейшего увеличения амплитуды сокращения. Такая реакция скелетной мышцы обусловлена ее строением. Она состоит из множества мышечных волокон, имеющих различную возбудимость и, следовательно, вовлечение их в реакцию идет постепенно: на пороговую силу раздражителя реагируют мышечные волокна с самой высокой возбудимостью, т.е. имеющие самый низкий порог раздражения. По мере увеличения силы раздражителя в сократительный процесс постепенно вовлекаются волокна, имеющие меньшую возбудимость. При максимальной силе раздражителя происходит сокращение всех мышечных волокон, составляющих данную мышцу, и поэтому амплитуда сокращений мышцы больше не увеличивается, несмотря на увеличение силы раздражителя.
Ход работы . Готовят препарат икроножной мышцы лягушки. Находят порог раздражения для мышцы, который определяют по ее минимальному сокращению. Далее, увеличивая силу раздражителя, записывают сокращение мышцы на кимографе.
Сделать вывод о зависимости между величиной раздражения и силой сокращения мышц.
6. Действие раздражителей различной природы.
Задача № 1.
При ухудшении кровоснабжения миокарда в межклеточной жидкости повышается концентрация ионов калия. Как и почему это скажется на генерации ПД в клетках миокарда?
Эталон ответа.
При повышении концентрации ионов калия в межклеточной жидкости возникает гиперполяризация мембран волокон миокарда. Значение их критического уровня деполяризации приближается к нулю, в результате чего генерация потенциала действия (ПД) станет невозможна.
Задача № 2.
Как и почему изменится амплитуда ПД клетки
а) при повышении концентрации ионов калия в цитоплазме
б) при повышении концентрации ионов натрия в межклеточной жидкости
в) при увеличении проницаемости мембраны клеток для ионов калия?
Эталон ответа.
Амплитуда ПД при повышении концентрации ионов калия в цитоплазме и увеличении проницаемости клеточной мембраны для этих ионов будет уменьшаться, а при повышении концентрации ионов натрия в межклеточной жидкости будет увеличиваться.
Задача № 3.
Какое практическое значение имеет следствие закона «силы-времени», согласно которому при предельно коротком времени действия сверхсильного раздражителя в ткани не будет возникать возбуждение?
Эталон ответа.
Данное свойство (закон, следствие закона «силы-времени») является биофизической основой метода УВЧ-терапии. Такой электрический ток вследствие своей ультравысокой частоты не успевает вызвать изменение состояния белков каналов и насосов мембран клеток, следовательно, генерации ПД в мышечных клетках и нервных окончаниях не происходит. Однако, вследствие наличия электрического сопротивления тканей происходит их нагревание.
Задача № 4.
Под влиянием местного анестетика в мембране клетки увеличилось число инактивированных натриевых каналов. Как и почему это скажется на параметрах ПД, возникающих в клетке?
Эталон ответа.
При увеличении числа инактивированных натриевых каналов клеточной мембраны уменьшится ее проводимость для ионов натрия. В результате этого уменьшится диффузионный поток положительно заряженных ионов натрия, входящий в клетку во время восходящей фазы ПД. Это приведет к уменьшению крутизны этой фазы и к уменьшению амплитуды ПД.
Задача № 5.
Под влиянием фармакологических факторов в мембране клеток увеличилось число калиевых каналов, которые могут активироваться при генерации ПД клетки. Как и почему это скажется на параметрах ПД клетки?
Эталон ответа.
Если во время генерации ПД увеличится число активированных калиевых каналов клеточной мембраны, то возрастет диффузионный поток положительно заряженных ионов калия, который выходит из клетки, в основном во время нисходящей фазы ПД. Это приведет к уменьшению длительности этой фазы, а, следовательно, и всего ПД в целом. Кроме того, может также несколько уменьшиться амплитуда ПД.
ЗАНЯТИЕ № 2
ТЕМА: ФИЗИОЛОГИЯ МЫШЦ
По морфологическим признакам выделяют три группы мышц:
1) поперечно-полосатые мышцы (скелетные мышцы);
2) гладкие мышцы;
3) сердечную мышцу (или миокард).
Функции поперечно-полосатых мышц:
1) двигательная (динамическая и статическая);
2) обеспечения дыхания;
3) мимическая;
4) рецепторная;
5) депонирующая;
6) терморегуляторная.
Функции гладких мышц:
1) поддержание давления в полых органах;
2) регуляция давления в кровеносных сосудах;
3) опорожнение полых органов и продвижение их содержимого.
Функция сердечной мышцы – насосная, обеспечение движения крови по сосудам.
1) возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала);
2) низкая проводимость, порядка 10–13 м/с;
3) рефрактерность (занимает по времени больший отрезок, чем у нервного волокна);
4) лабильность;
5) сократимость (способность укорачиваться или развивать напряжение).
Различают два вида сокращения:
а) изотоническое сокращение (изменяется длина, тонус не меняется);
б) изометрическое сокращение (изменяется тонус без изменения длины волокна). Различают одиночные и титанические сокращения. Одиночные сокращения возникают при действии одиночного раздражения, а титанические возникают в ответ на серию нервных импульсов;
6) эластичность (способность развивать напряжение при растягивании).
Физиологические особенности гладких мышц.
Гладкие мышцы имеют те же физиологические свойства, что и скелетные мышцы, но имеют и свои особенности:
1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса;
2) самопроизвольную автоматическую активность;
3) сокращение в ответ на растяжение;
4) пластичность (уменьшение растяжения при увеличении растяжения);
5) высокую чувствительность к химическим веществам.
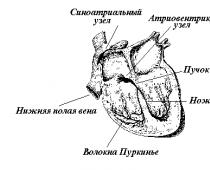
Физиологической особенностью сердечной мышцы является ее автоматизм . Возбуждение возникает периодически под влиянием процессов, протекающих в самой мышце. Способностью к автоматизму обладают определенные атипические мышечные участки миокарда, бедные миофибриллами и богатые саркоплазмой.
2. Механизмы мышечного сокращения
Электрохимический этап мышечного сокращения.
1. Генерация потенциала действия. Передача возбуждения на мышечное волокно происходит с помощью ацетилхолина. Взаимодействие ацетилхолина (АХ) с холинорецепторами приводит к их активации и появлению потенциала действия, что является первым этапом мышечного сокращения.
2. Распространение потенциала действия. Потенциал действия распространяется внутрь мышечного волокна по поперечной системе трубочек, которая является связывающим звеном между поверхностной мембраной и сократительным аппаратом мышечного волокна.
3. Электрическая стимуляция места контакта приводит к активации фермента и образованию инозилтрифосфата, который активирует кальциевые каналы мембран, что приводит к выходу ионов Ca и повышению их внутриклеточной концентрации.
Хемомеханический этап мышечного сокращения.
Теория хемомеханического этапа мышечного сокращения была разработана О. Хаксли в 1954 г. и дополнена в 1963 г. М. Девисом. Основные положения этой теории:
1) ионы Ca запускают механизм мышечного сокращения;
2) за счет ионов Ca происходит скольжение тонких актиновых нитей по отношению к миозиновым.
В покое, когда ионов Ca мало, скольжения не происходит, потому что этому препятствуют молекулы тропонина и отрицательно заряды АТФ, АТФ-азы и АДФ. Повышенная концентрация ионов Ca происходит за счет поступления его из межфибриллярного пространства. При этом происходит ряд реакций с участием ионов Ca:
1) Ca2+ реагирует с трипонином;
2) Ca2+ активирует АТФ-азу;
3) Ca2+ снимает заряды с АДФ, АТФ, АТФ-азы.
Взаимодействие ионов Ca с тропонином приводит к изменению расположения последнего на актиновой нити, открываются активные центры тонкой протофибриллы. За счет них формируются поперечные мостики между актином и миозином, которые перемещают актиновую нить в промежутки между миозиновой нитью. При перемещении актиновой нити относительно миозиновой происходит сокращение мышечной ткани.
Итак, главную роль в механизме мышечного сокращения играют белок тропонин, который закрывает активные центры тонкой протофибриллы и ионы Ca.
Физиология скелетных и гладких мышц
Лекция 5
У позвоночных и человека три вида мышц : поперечнополосатые мышцы скелета, поперечнополосатая мышца сердца – миокард и гладкие мышцы, образуюцие стенки полых внутренних органов и сосудов.
Анатомической и функциональной единицей скелетных мышц является нейромоторная единица - двигательный нейрон и иннервируемая им группа мышечных волокон. Импульсы, посылаемые мотонейроном, приводят в действие все образующие ее мышечные волокна.
Скелетные мышцы состоят из большого количества мышечных волокон. Волокно поперечнополосатой мышцы имеет вытянутую форму, диаметр его от 10 до 100 мкм, длина волокна от нескольких сантиметров до 10-12 см. Мышечная клетка окружена тонкой мембраной – сарколеммой , содержит саркоплазму (протоплазму) и многочисленные ядра . Сократительной частью мышечного волокна являются длинные мышечные нити – миофибриллы , состоящие в основном из актина, проходящие внутри волокна от одного конца до другого, имеющие поперечную исчерченность. Миозин в гладких мышечных клетках находится в дисперсном состоянии, но содержит много белка, играющего важную роль в поддержании длительного тонического сокращения.
В период относительного покоя скелетные мышцы полностью не расслабляются и сохраняют умеренную степень напряжения, т.е. мышечный тонус .
Основные функции мышечной ткани:
1)двигательная – обеспечение движения
2)статическая – обеспечение фиксации, в том числе и в определенной позе
3)рецепторная – в мышцах имеются рецепторы, позволяющие воспринимать собственные движения
4)депонирующая – в мышцах запасаются вода и некоторые питательные вещества.
Физиологические свойства скелетных мышц:
Возбудимость . Ниже, чем возбудимость нервной ткани. Возбуждение распространяется вдоль мышечного волокна.
Проводимость . Меньше проводимости нервной ткани.
Рефрактерный период мышечной ткани более продолжителен, чем нервной ткани.
Лабильность мышечной ткани значительно ниже, чем нервной.
Сократимость – способность мышечного волокна изменять свою длину и степень напряжения в ответ на раздражение пороговой силы.
При изотоническом сокращении изменяется длина мышечного волокна без изменения тонуса. При изометрическом сокращении возрастает напряжение мышечного волокна без изменения его длины.
В зависимости от условий стимуляции и функционального состояния мышцы может возникнуть одиночное, слитное (тетаническое) сокращение или контрактура мышцы.
Одиночное мышечное сокращение. При раздражении мышцы одиночным импульсом тока возникает одиночное мышечное сокращение.
Амплитуда одиночного сокращения мышцы зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных групп волокон различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна. При увеличении силы раздражающего тока в процесс возбуждения вовлекаются и менее возбудимые группы мышечных волокон; амплитуда сокращений суммируется и растет до тех пор, пока в мышце не останется волокон, не охваченных процессом возбуждения. В этом случае регистрируется максимальная амплитуда сокращения, которая не увеличивается, несмотря на дальнейшее нарастание силы раздражающего тока.
Тетаническое сокращение . В естественных условиях к мышечным волокнам поступают не одиночные, а ряд нервных импульсов, на которые мышца отвечает длительным, тетаническим сокращением, или тетанусом . К тетаническому сокращению способны только скелетные мышцы. Гладкие мышцы и поперечнополосатая мышца сердца не способны к тетаническому сокращению из-за продолжительного рефрактерного периода.
Тетанус возникает вследствие суммации одиночных мышечных сокращений. Чтобы возник тетанус, необходимо действие повторных раздражений (или нервных импульсов) на мышцу еще до того, как закончится ее одиночное сокращение.
Если раздражающие импульсы сближены и каждый из них приходится на тот момент, когда мышца только начала расслабляться, но не успела еще полностью расслабиться, то возникает зубчатый тип сокращения (зубчатый тетанус ).
Если раздражающие импульсы сближены настолько, что каждый последующий приходится на время, когда мышца еще не успела перейти к расслаблению от предыдущего раздражения, то есть происходит на высоте ее сокращения, то возникает длительное непрерывное сокращение, получившее название гладкого тетануса .
Гладкий тетанус – нормальное рабочее состояние скелетных мышц обусловливается поступлением из ЦНС нервных импульсов с частотой 40-50 в 1с.
Зубчатый тетанус возникает при частоте нервных импульсов до 30 в 1с. Если мышца получает 10-20 нервных импульсов в 1с, то она находится в состоянии мышечного тонуса , т.е. умеренной степени напряжения.
Утомление мышц . При длительном ритмическом раздражении в мышце развивается утомление. Признаками его являются снижение амплитуды сокращений, увеличение их латентных периодов, удлинение фазы расслабления и, наконец, отсутствие сокращений при продолжающемся раздражении.
Еще одна разновидность длительного сокращения мышц - контрактура . Она продолжается и при снятии раздражителя. Контрактура мышцы наступает при нарушении обмена веществ или изменении свойств сократительных белков мышечной ткани. Причинами контрактуры могут быть отравление некоторыми ядами и лекарственными средствами, нарушение обмена веществ, повышение температуры тела и другие факторы, приводящие к необратимым изменениям белков мышечной ткани.
По структуре гладкая мышца отличается от поперечнополосатой скелетной мышцы и мышцы сердца. Она состоит из клеток веретенообразной формы длиной от 10 до 500 мкм, шириной 5-10 мкм, содержащих одно ядро. Гладкомышечные клетки лежат в виде параллельно ориентированных пучков, расстояние между ними заполнено коллагеновыми и эластическими волокнами, фибробластами, питающими магистралями. Мембраны прилежащих клеток образуют нексусы, которые обеспечивают электрическую связь между клетками и служат для передачи возбуждения с клетки на клетку. Кроме того плазматическая мембрана гладкомышечной клетки имеет особые впячивания - кавеолы, благодаря которым площадь мембраны увеличивается на 70%. Снаружи плазматическая мембрана покрыта базальной мембраной. Комплекс базальной и плазматической мембраны называют сарколеммой. В гладкой мышцы отсутствуют саркомеры. Основу сократительного аппарата составляют миозиновые и актиновые протофибриллы. В ГМК актиновых протофибрилл намного больше, чем в поперечно-полосатом мышечном волокне. Соотношение актин/миозин = 5:1.
Толстые и тонкие миофиламеты распылены по всей саркоплазме гладкого миоцита и не имеют такой стройной организации, как в поперечно-полосатой скелетной мышце. При этом тонкие филаменты прикрепляются к плотным тельцам. Некоторые из этих телец расположены на внутренней поверхности сарколеммы, но большинство из них находятся в саркоплазмме. Плотные тельца состоят из альфа-актинина – белка обнаруженного в структуре Z-мембраны поперечнополосатых мышечных волокон. Некоторые из плотных телец расположенных на внутренней поверхности мембраны соприкасаются с плотными тельцами прилегающей клетки. Тем самым сила, создаваемая одной клеткой может передаваться следующей. Толстые миофиламенты гладкой мышцы содержат миозин, а тонкие – актин, тропомиозин. При этом в составе тонких миофиламентов не обнаружен тропонин.
Гладкие мышцы встречаются в стенках кровеносных сосудах, коже и внутренних органах.
Гладкая мышца играет важную роль в регуляции
просвета воздухоносных путей,
тонуса кровеносных сосудов,
двигательной активности желудочнокишечного тракта,
матки и др.
Классификация гладких мышц:
Мультиунитарные, входят в состав цилиарной мышцы, мышц радужки глаза, мышцы поднимающей волос.
Унитарные (висцеральная), находятся во всех внутренних органах, протоках пищеварительных желез, кровеносных и лимфатических сосудах, коже.
Мультиунитарная гладкая мышца.
состоит из отдельных гладкомышечных клеток, каждая из которых, находится независимо друг от друга;
имеет большую плотность иннервации;
как и поперечнополосатые мышечные волокна, снаружи покрыты веществом, напоминающим базальную мембрану, в состав которого входят, изолирующие клетки друг от друга, коллагеновые и гликопротеиновые волокна;
каждая мышечная клетка может сокращаться отдельно и ее активность регулируется нервными импульсами;
Унитарная гладкая мышца (висцеральная).
представляет собой пласт или пучок, а сарколеммы отдельных миоцитов имеют множественные точки соприкосновения. Это позволяет возбуждению распространяться от одной клетки к другой
мембраны рядом расположенных клеток образуют множественные плотные контакты (gap junctions), через которые ионы имеют возможность свободно передвигаться из одной клетки в другу
потенциал действия, возникающий на мембране гладкомышечной клетки, и ионные потоки могут распространяться по мышечному волокну, обеспечивая возможность одновременного сокращения большого количества отдельных клеток. Данный тип взаимодействия известен как функциональный синцитий
Важной особенность гладкомышечных клеток является их способность к самовозбуждению (автоматии) , то есть они способны генерировать потенциал действия без воздействия внешнего раздражителя.
Постоянный мембранный потенциал покоя в гладких мышцах отсутствует, он постоянно дрейфует и в среднем составляет -50мВ. Дрейф происходит спонтанно, без каких-либо влияний и когда мембранный потенциал покоя достигает критического уровня возникает потенциал действия, который и вызывает сокращение мышцы. Продолжительность потенциала действия достигает нескольких секунд, поэтому и сокращение тоже может длиться несколько секунд. Возникшее возбуждение затем распространяется через нексус на соседние участки вызывая их сокращения.
Спонтанная (независимая) активность связана с растяжением гладкомышечных клеток и когда они растягиваются возникает потенциал действия. Частота возникновения потенциалов действия зависит от степени растяжения волокна. Например, перистальтические сокращения кишечника усиливаются при растягивании его стенок химусом.
Унитарные мышцы в основном сокращаются под влиянием нервных импульсов, но иногда возможны и спонтанные сокращения. Одиночный нервный импульс не способен вызывать ответной реакции. Для ее возникновение необходимо суммировать несколько импульсов.
Для всех гладких мышц при генерации возбуждения характерна активация кальциевых каналов, поэтому в гладких мышцах все процессы идут медленнее по сравнению со скелетной.
Скорость проведения возбуждения по нервным волокнам к гладким мышцам составляет 3-5 см в секунду.
Одним из важных раздражителей инициирующих сокращение гладких мышц является их растяжение. Достаточное растяжение гладкой мышцы обычно сопровождается появлением потенциалов действия. Таким образом, появлению потенциалов действия при растяжении гладкой мышцы способствует два фактора:
медленные волновые колебания мембранного потенциала;
деполяризация, вызываемая растяжением гладкой мышцы.
Данное свойство гладкой мышцы позволяет ей автоматически сокращаться при растяжении. Например, во время переполнения тонкого кишечника возникает перистальтическая волна, которая и продвигает содержимое.
Сокращение гладкой мышцы.
Гладкие мышцы, как и поперечно-полосатые, содержат миозин, с поперечными мостиками, гидролизующий АТФ, и для того, чтобы вызвать сокращение, взаимодействует с актином. В противоположность поперечно-полосатым мышцам, тонкие филаменты гладких мышц содержат только актин и тропомиозин и не содержат тропонин; регуляция сократительной активности в гладких мышцах происходит благодаря связыванию Са ++ с кальмодулином, активирующим киназу миозина, которая фосфорилирует регуляторную цепь миозина. Это приводит к гидролизу АТФ и запускает цикл образования поперечных мостиков. В гладкой мышце движение актомиозиновых мостиков является более медленным процессом. Распад молекул АТФ и высвобождение энергии, необходимой для обеспечения движения актомиозиновых мостиков происходит не так быстро как в поперечнополосатой мышечной ткани.
Экономичность энергозатрат в гладкой мышце является чрезвычайно важным в общем потреблении организмом энергии, так как, кровеносные сосуды, тонкий кишечник, мочевой пузырь, желчный пузырь и другие внутренние органы постоянно находятся в тонусе.
Во время сокращения гладкая мышца способна укорачиваться вплоть до 2/3 ее первоначальной длины (скелетная мышца от 1/4 до 1/3 длины). Это позволяет полым органам выполнять свою функцию изменяя свой просвет в значительных пределах.